蛋白質(zhì)被泛素-蛋白酶體系統(tǒng)降解需經(jīng)過以下過程:①靶蛋白的泛素化;②靶蛋白的降解。三種重要的酶共同介導(dǎo)了靶蛋白泛素化過程:泛素活化酶(ubiquitin activating enzyme,E1)、泛素結(jié)合酶(ubiquitin conjugating enzyme,E2)和泛素連接酶(ubiquitin-protein ligase,E3)。E1水解ATP后與泛素分子C端形成硫酯鍵,然后E1將活化的泛素遞交給E2,最后泛素在E3的募集作用下,從E2轉(zhuǎn)移到靶蛋白上,或者靶蛋白首先與E3結(jié)合,泛素再轉(zhuǎn)移過來。泛素化的靶蛋白最終進(jìn)入26S蛋白酶體而被降解,而泛素分子則從泛素鏈中分離開來,從而實(shí)現(xiàn)重復(fù)利用。關(guān)于E1的作用機(jī)制及具體數(shù)目尚不明確,在人類組織中至少已發(fā)現(xiàn)了40多種E2及700多種E3。且E3具有高度的底物特異性,通過識別和結(jié)合特異的靶蛋白序列或降解決定子(degron),即N末端法則實(shí)現(xiàn)對靶蛋白的選擇性降解。
泛素一蛋白酶體途徑(ubiquitin-proteasome pathway)是細(xì)胞內(nèi)一個(gè)重要的蛋白質(zhì)降解調(diào)節(jié)系統(tǒng)。通過對底物蛋白的多聚泛素化并經(jīng)蛋白酶體降解,可以影響或調(diào)節(jié)多種細(xì)胞活動(dòng),包括:基因轉(zhuǎn)錄、細(xì)胞周期調(diào)節(jié)、免疫反應(yīng)、細(xì)胞受體功能及腫瘤生長、炎癥過程等。該途徑也是一種動(dòng)態(tài)的蛋白質(zhì)雙向修飾調(diào)控系統(tǒng),在體內(nèi)由泛素連接酶系統(tǒng)(E1-E2-E3)對底物進(jìn)行泛素化修飾,去泛素化酶(DUB)家族負(fù)責(zé)通過水解泛素羧基末端的酯鍵、肽鍵或異肽鍵,將泛素分子特異性的從鏈接有泛素的蛋白質(zhì)或者前體蛋白水解下來,起到去泛素化的作用,對蛋白降解進(jìn)行反向調(diào)節(jié),從而影響蛋白質(zhì)的功能。

去泛素化酶是一類數(shù)量很大的蛋白酶家族,主要分為五個(gè)家族,分別是泛素羧基末端水解酶(UCH)家族,泛素特異性蛋白酶(USP/UBP)家族,Otubaim(OTU)家族,Josephin結(jié)構(gòu)域蛋白家族及JAMM家族。
在細(xì)胞內(nèi)去泛素化酶的功能可大致分為以下幾種: (1)加工泛素前體, 產(chǎn)生自由的泛素分子; (2)移除蛋白質(zhì)上的泛素鏈, 避免蛋白質(zhì)被蛋白酶體降解, 從而穩(wěn)定蛋白質(zhì); (3) 移除蛋白質(zhì)上連接的非降解泛素化信號; (4)通過阻 止泛素分子與底物蛋白質(zhì)一起被降解, 確保細(xì)胞內(nèi)泛素分子的穩(wěn)態(tài); (5)參與細(xì)胞內(nèi)游離泛素鏈的解體; (6)通過剪切泛素鏈, 編輯泛素鏈的類型。

去泛素化酶(Deubiquitinating enzymes, DUB):與泛素化修飾需要E1、E2及E3三種酶共同作用不同,去泛素化的過程只需要DUBs,且DUBs作用的底物蛋白是被泛素修飾的蛋白(圖1)。DUBs通過水解泛素分子羧基末端的肽鍵、異肽鍵或酯鍵從而去除底物蛋白中的泛素或通過縮短共軛泛素鏈來抵消泛素化,進(jìn)而抑制底物蛋白的降解(Vogel K and Isono E., 2024)。
圖1 泛素化和去泛素化過程(Zheng et al., 2023)。泛素化過程由E1泛素活化酶、E2泛素偶聯(lián)酶和E3泛素連接酶完成。E1酶激活泛素并將其轉(zhuǎn)移到E2酶。然后,E2酶與E3酶相互作用,導(dǎo)致泛素從E2酶轉(zhuǎn)移到特定的靶蛋白。泛素化蛋白可以被蛋白酶體識別和降解。泛素化可以通過去泛素化逆轉(zhuǎn),去泛素化是一種將泛素從靶蛋白中分離出來的過程。
去泛素化酶的分類
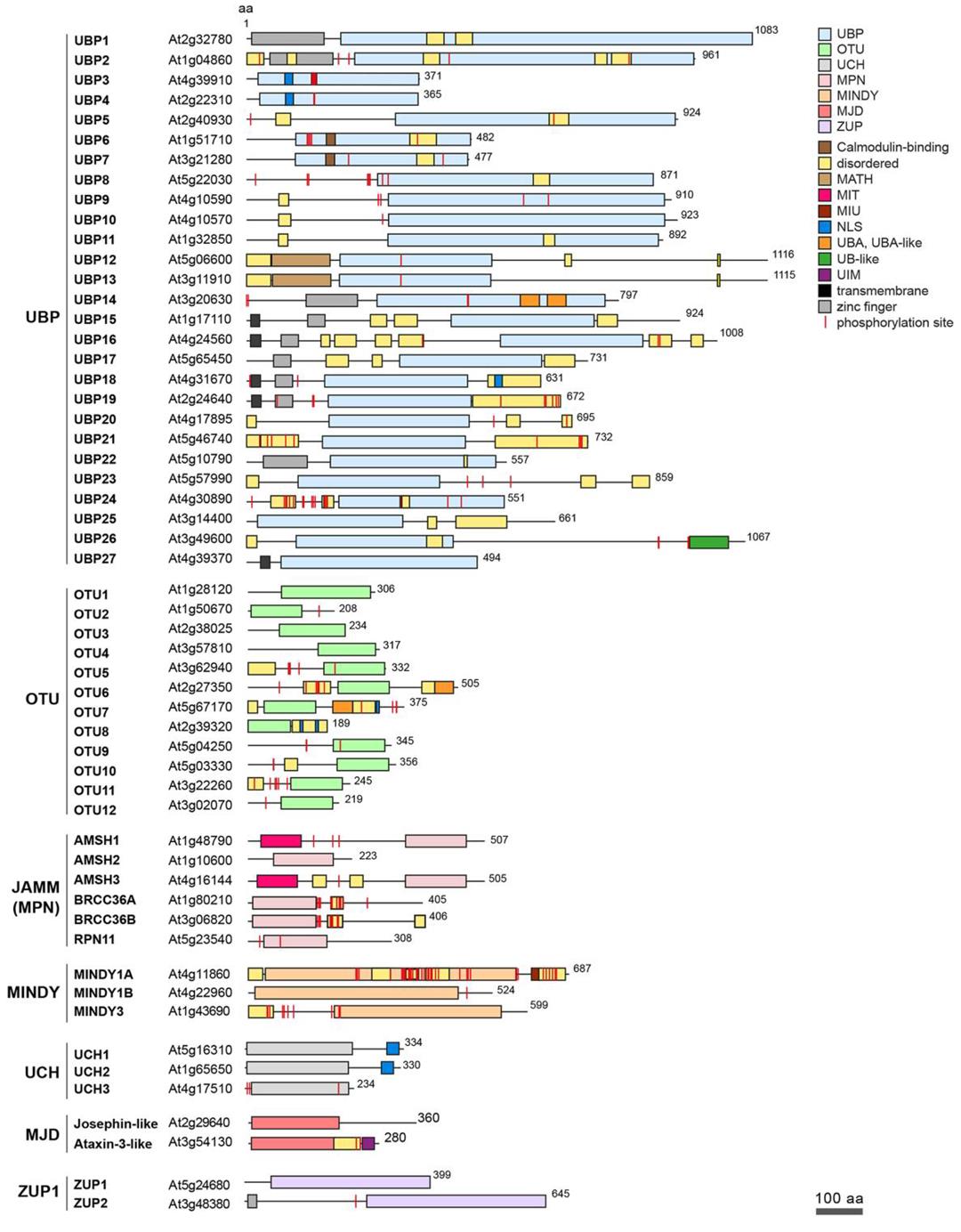
在真核生物中,根據(jù)催化機(jī)制的不同,DUBs可分為兩大類:半胱氨酸蛋白酶家族和金屬蛋白酶家族(圖2)。其中,半胱氨酸蛋白酶家族包括泛素特異性蛋白酶(USPs)、泛素C端水解酶(UCHs)、Machado?Joseph結(jié)構(gòu)域蛋白酶(MJDs)、卵巢腫瘤蛋白酶(OUTs)、Motif與泛素分子結(jié)合的DUB家族蛋白酶(MINDYs)和含鋅指泛素肽酶(ZUP/ZUFSP)六類;而金屬蛋白酶家族只包含MPN(+)/JAMM蛋白酶家族(Clague et al., 2019)。根據(jù)當(dāng)前已有的DUBs研究結(jié)果, Vogel K和Isono E兩位研究者統(tǒng)計(jì)了擬南芥中DUBs的種類及其對應(yīng)的結(jié)構(gòu)域(圖3)。
圖2 特異性DUBs的催化機(jī)制(Vogel K and Isono E., 2024)。(A)以泛素二聚體為底物的半胱氨酸蛋白酶DUBs的催化機(jī)制。S1位點(diǎn)結(jié)合末端或遠(yuǎn)端泛素(UB環(huán)),而S1'位點(diǎn)結(jié)合近端泛素,該泛素有一個(gè)游離的C端或與靶蛋白結(jié)合。這些結(jié)合位點(diǎn)有助于將遠(yuǎn)端泛素的C端甘氨酸(G)和近端泛素的賴氨酸(K)之間的異肽鍵定位在活性位點(diǎn)附近。活性位點(diǎn)三聯(lián)體由半胱氨酸(C)、組氨酸(H)和天冬氨酸(D)組成。在第一步中,半胱氨酸側(cè)鏈的硫陰離子在異肽鍵的親電碳原子上發(fā)生親核攻擊。半胱氨酸側(cè)鏈的硫陰離子被質(zhì)子化組氨酸側(cè)鏈穩(wěn)定,而組氨酸側(cè)鏈又被天冬氨酸側(cè)鏈穩(wěn)定。親核攻擊后,催化位點(diǎn)半胱氨酸的硫原子與異肽鍵的碳原子之間形成臨時(shí)鍵,導(dǎo)致異肽鍵斷裂,近端泛素釋放。親電的碳原子是硫酯鍵的一部分,可以被來自水分子的親核氧攻擊。隨后,來自水的羥基與遠(yuǎn)端泛素的C端甘氨酸的碳原子結(jié)合,將遠(yuǎn)端泛素從催化側(cè)移除。催化位點(diǎn)組氨酸被來自水分子的第二個(gè)質(zhì)子質(zhì)子化;(B)金屬蛋白酶DUBs的催化機(jī)理。遠(yuǎn)端泛素和近端泛素分別與S1和S1'位點(diǎn)結(jié)合,使異肽鍵靠近活性位點(diǎn)。活性位點(diǎn)包括一個(gè)絲氨酸(S)、一個(gè)天冬氨酸(D)、兩個(gè)組氨酸(H)、一個(gè)谷氨酸(E)和一個(gè)由組氨酸和天冬氨酸配位的鋅質(zhì)子(Zn2+)。在泛素結(jié)合之前,活性位點(diǎn)的鋅質(zhì)子與水分子發(fā)生反應(yīng),這使得水分子處于活性中心,允許它的1個(gè)質(zhì)子與谷氨酸側(cè)鏈上帶負(fù)電的氧原子結(jié)合,進(jìn)而形成一個(gè)帶負(fù)電荷的羥基,這個(gè)羥基此時(shí)處于攻擊異肽鍵的親電碳的最佳位置。當(dāng)與泛素結(jié)合時(shí),來自異肽鍵的親電碳受到來自水的親核氧的攻擊,該反應(yīng)通過活性位點(diǎn)絲氨酸對異肽鍵羰基氧原子的暫時(shí)穩(wěn)定促進(jìn)。水的親核攻擊引起異肽鍵的水解,導(dǎo)致遠(yuǎn)端和近端泛素的釋放。遠(yuǎn)端泛素的C末端甘氨酸與來自水分子的羥基結(jié)合,而近端泛素賴氨酸側(cè)鏈的氨基則與來自水分子的質(zhì)子結(jié)合。參與反應(yīng)的側(cè)鏈官能團(tuán)用骨架式表示,共價(jià)鍵用黑色實(shí)線表示,非共價(jià)鍵用黑色虛線表示。
圖3 擬南芥DUBs的種類及結(jié)構(gòu)域(Vogel K and Isono E., 2024)。

UbiQBio成立于2010年,位于荷蘭阿姆斯特丹,專注于泛素(ubiquitin)及其類似蛋白的研究工具開發(fā)。公司由教授Huib Ovaa、Farid El Oualid和Alfred Nijkerk創(chuàng)立,致力于為藥物發(fā)現(xiàn)。技術(shù)平臺由三項(xiàng)專有技術(shù)組成: Ubi0Bl0ckm.Ubi0-Cick" 和 Ubi0-Swn™,這使我們能夠開發(fā)泛素 (Ub)領(lǐng)域所缺乏的產(chǎn)品和服務(wù),利用這些化學(xué)合成技術(shù),我們可以構(gòu)建件何U6綴合物、Ub突變體及其組合。現(xiàn)在可以更有效地開發(fā)已知試劑,但最重要的是構(gòu)建遠(yuǎn)遠(yuǎn)超出任何當(dāng)前可用(生物)方法所能達(dá)到的試劑。借助我們的專有技術(shù)平臺,我們能夠使用自動(dòng)同相肽合成技術(shù)合成天然構(gòu)象的位點(diǎn)選擇性泛素化肽。
靶點(diǎn)科技(北京)有限公司
地址:中關(guān)村生命科學(xué)園北清創(chuàng)意園2-4樓2層
© 2025 版權(quán)所有:靶點(diǎn)科技(北京)有限公司 備案號:京ICP備18027329號-2 總訪問量:337856 站點(diǎn)地圖 技術(shù)支持:化工儀器網(wǎng) 管理登陸